
The physiological complexity of the fungi is manifest in their diversity; there are estimated to be 1.5 million different species of which only about 69,000 have been identified. These statistics are indicative of the reproductive fecundity of the fungi in initiating and replicating a broad taxonomy and of the limitations in our understanding of their nature, which includes the physiology of the reproductive process. While the reproductive processes of the Kingdoms Plantae and Animalia are well known, the fungi of the Kingdom Eumycota (also known as Myceteae), are not. Sex refers to the division of living species according to their reproductive roles, traditionally the female as the guardian of the egg and the male as the purveyor of the sperm. This dichotomy is very distinct in animals and to a somewhat lesser extent plants; in a perfect flower both sexes are represented. Some fungi, particularly the simpler and more primitive forms, have something more or less analogous to male and female sexes. However, the more complicated forms like the basidiomycete mushrooms have a much more complex relationship that involves multiple combinations of paired individuals whose union results in reproduction. In order to comprehend mushroom reproduction and attendant “sexuality”, it is necessary to establish a lexical framework on which to base the discussion.
The most important aspect of reproduction is the transmission of the DNA code from one generation to the next, as it critically establishes speciation and allows for random mutation. Reproduction is initiated by the union of the male sperm and the female egg in most animals, the pollen from the male anther and female pistil in (flowering) plants, and by several variations on the theme; the end result is the same. Both males and females carry the same number of chromosomes; the number varies according to species (23 in humans) and is usually denoted simply by the letter n; the single chromosome set cells are called haploid, derived from the Greek word for ‘single’. When the two haploids combine, the result is 2n; a fertilized egg or zygote – also a double haploid or diploid. The zygote, as the only repository for all of the genetic information of the new organism, must grow without losing the code. This is accomplished by mitosis, the division of the nucleus into two identical daughter cells which have the same diploid 2n genetic makeup. The word was derived from the Greek mitos meaning thread and the Latin osis meaning process based on observations of the thread-like chromosomes in the nuclei. As the diploid nuclei continue to divide, the resultant cells differentiate according to the established sequence of gene expression, the eventual result a fully grown individual species. Reproduction of a new individual requires that the diploid 2n cell be reduced to a haploid n cell in order for it to seek out a haploid cell of the opposite sex. The reduction of a 2n diploid to an n haploid was termed meiosis, from the Greek meion meaning less and osis, meaning process; since this is a homonym for mitosis, differentiating them is the pons asinorum of biology students. With refinements in laboratory methods and instrumentation in the late 19th Century, it was discovered that meiosis was a two-step process so that four haploid cells were formed. In summary, two n or haploid nuclei combine to produce a 2n diploid which grows from one cell to many with mitosis; the 2n diploid cell reduces to four n haploid cells by meiosis to complete the cycle.
The fungi can be divided into two generalized groups based on their reproductive strategies: sexual and asexual. In the mycological vernacular, sexual fungi are sometimes referred to as the Fungi Perfecti and asexual as (naturally) Fungi Imperfecti. On also occasionally finds the asexual fungi referred to as Deuteromyctes from the Greek word deuesthai, ‘to be in need of’ and mycetes or fungi, the etymological suggestion is that to be asexual is to need sex. More properly, the sexual fungus is called a teleomorph and the asexual fungus is called an anamorph. This also makes good etymological sense since teleo is the Greek word for ‘consummation’ or ‘completeness’ whereas ana means simply ‘on’ or ‘up’, implying simplicity. Both teleomorphs and anamorphs produce spores to execute the reproductive function. The sexual spores are appropriately called teleospores; they convey a genetic component from two separate parent entities (which really should not be called male and female). Asexual spores are not called anaspores, though they probably should be if language were based on logic, but rather mitospores, to indicate that they are the product of mitotic or asexual division. Another name that one encounters for asexual spores is conidium, which comes from the Greek word konis, meaning dust; these are asexual spores that are formed outside any enclosing structure. Mushroom sex is complicated.
The anamorphs largely occupy the netherworld of the Kingdom Eumycota, which is taxonomically comprised of three phyla. The primitive Phylum Chytridiomycota (one species of chytrid is responsible for the recent decimation of many amphibian species in Central America and Australia – the frog die-off) is comprised of species which have asexual zoospores that move with a whip-like flagellum. The Phylum Zygomycota is a complex group that includes the bread molds and the soil fungi that are the predominant in the establishing mycorrhizal relationships with plant roots. Zygomycetes propagate by both teleospores and mitospores; they are classified by the unique sexual phase that results from a physical conjugation (the Greek word for union is zygoma) of two separate fungi. However, it is the third and most physically obvious Phylum Dikaryomycota which is comprised of the cup-like Subphylum Ascomycotina and the true mushroom Subphylum Basidiomycotina that have the more confusingly exasperating reproductive systems. The ascomycetes and the basidiomycetes can have both an anamorph and a teleomorph, in which case the two taken together may be called a holomorph. Some species exist only as anamorphs, some (so far as is known) exist only as teleomorphs, and some have part of the life cycle as anamorph and part as teleomorph. Deciphering the complexities of the holomorph which can involve multiple hosts and multiple physical forms is a seemingly Sisyphean task, though it has been worked out for a few species – where there is some agronomic significance. A case in point is the basidiomycete species Puccinia graminis subspecies tritici, more commonly known as wheat rust. The life cycle starts when a teleospore germinates on a barberry (Berberis), a popular garden shrub (the Japanese barberry, B. thunbergii has escaped cultivation and become an invasive species) to form a brown pustule that doesn’t harm the barberry, but which emits sexual spores that germinate on wheat plants (Triticum) and form anamorphs that create more spores that can cause massive wheat crop damage by investing acres of (amber waves of) grain. During the late summer, the fungus shifts to the production of teleospores that are dispersed in search of the barberry. Because of this, a barberry eradication program was initiated by the U. S Department of Agriculture in 1918 that continued until 1975, destroying an estimated 100 million plants. There are likely thousands of holomorphs (an anamorph and a teleomorph) that have yet to be fully characterized, if even known to exist. The fungal world is complex.
Since anamorphs are asexual, there is no question about sex; there is none. When a mitospore comes to rest in a provident environment, it germinates to create a filamentous tendril called a hypha (from the Greek hyphos, meaning web) that grows in what is called the assimilative mode. Assimilation is the process by which food is changed from the form in which it is found into the form in which it can be used; it is the fungal equivalent of plant vegetative growth. As assimilative growth continues, the bundles of hyphae eventually form a tangled “web” called a mycelium, which comes from a combination of Greek words that translate loosely as “folded warty fungus.” In the case of the fungi, assimilation is most generally in the form of the secretion of enzymes from the hyphal tip that break down the plant or animal nutrients into a form that can be used by the fungus for growth. The hypha extends, becoming ramose to exploit new resources, until the host is invested and growth ceases. Reproduction occurs when there is sufficient energy available for the production of spores, and, most importantly, when the food source is depleted. The process of spore formation among anamorphs is generally straight forward. In the simplest case, structures called conidiophores grow directly on the hypha and produce conidia, the mitospores of the anamorph. In some cases, a simple, specialized spore bearing body called a sporangium will be created. In either case, prodigious quantities of spores are produced, and this is the key to the success of the fungi. The miniscule size of the spore permits airborne transmission over great distances and their ubiquity helps to ensure that at least one will be successful. For the asexual anamorphs, this is all that it takes. It is much harder for the sexual fungi. One may well posit that it makes no sense to invoke the complexities of sexuality when asexuality is so fecund. It is important to keep in mind that asexuality brooks very little, if any variation, whereas variation is the province and provenance of sexuality. Organisms can only evolve to changing environmental conditions efficiently and effectively only with the evolution that sexuality provides. And the environment is always changing.
Sexual reproduction in the dicaryomycotan fungi is simultaneously very simple and very complicated. According to Nicholas Money in Mr. Bloomfield’s Garden, “sexual behavior in mushroom-forming fungi spans monogamy and civility, to group sex and slaughter.” Bryce Kendrick in The Fifth Kingdom has a more nuanced view in noting that “reproduction in fungi frequently involves sex, though their sexual behavior is sometimes obscure.” The hidden sex of spore producing species has a turbid history; mosses, ferns, algae and fungi were at one point classified as in a subkingdom called Cryptogamia (literally hidden life). Before the invention of the microscope, spores were essentially invisible; the reproduction of any of the cryptogams was accordingly shrouded in the aura of thaumaturgic intrigue. For example, the absence of visible seeds in ferns led to some interesting interpretations as to the nature of fern propagation. As the fern was clearly a plant, then it must have a seed, and, by syllogistic logic, since the “fern seed” could not be seen, it was claimed by some early herbalists that it must be invisible. This ultimately led to a widely held belief that the invisibility of the fern seed conveyed invisibility to the bearer of the seed, but only if the seeds were collected at midnight on Midsummer Night’s Eve, June 23, also known as the eve of Saint John, the shortest night of the year. Mushrooms, whose reproductive mechanisms were even more obscure than those of the ferns, were seen as even more mysterious, a perspective enhanced by their seemingly chthonian appearance overnight, sometimes in fairy ring circles.

The simple part of mushroom sexuality is that they do not have any specialized sex organs. Any of the filamentous hyphae can engage in sexual union if approached with a sexually compatible hypha from another individual. The complicated part is that there may be many different pairing combinations that are sexually compatible, just as there will be many that are not. When the hyphae from two incompatible forms make contact, the result is a fungal battle for territory, the two strains defending their borders with melanin barriers; trees that are subject to fungal hegemony have patterned wood whose whorls and shadings afford an aesthetic effect employed by wood carvers. When the hyphae of two sexually compatible individuals make contact, the reproductive process begins. The complexity is in the breadth and extent of sexually compatibility. The multiple sexes of the basidiomycete fungi were first discovered by the German botanist Karl Johannes Kniep during the First World War through the evaluation of the fungus Schizophyllum commune, known as the common split gill. The choice of this fungus was not serendipitous; S. commune is distributed worldwide and grows throughout the year, a consequence of its high spore germination rate and its ability to thrive in a fairly broad range of environmental conditions. Kniep found that, unlike animals and plants that have one set of genes for each of the male and female genders, most fungi have two sets of genes which are called ‘mating type factors’; he called them A and B, a designation that has persisted. Over the last century, work with S. commune has revealed that there are about 340 A factors, and 64 B factors, which results in something like 21,000 possible “sexual” pairings. There have been estimates made for other fungi, but it should be recognized that the only way to determine if two types are compatible is to pair them to see what happens, a daunting and painstaking laboratory assignment. For most mushrooms, the number of mating factors, or sexes, is a matter of conjecture. It is likely, however, to be significantly more than two.
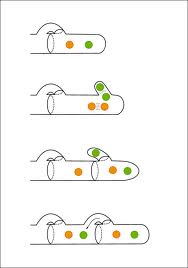
So, what happens when two hypha like each other? They mate. What this means for fungi is that the two hyphae merge to form a single cell; the union to form into a single cytoplasm is called plasmogamy. Since each of the two “sexes” are haploid (n), this results in two nuclei inside one cell (n + n). This is not the same as diploid (2n) because the nuclei remain distinct and separate. The cell with two different nuclei is called a dikaryon, from which the Phylum name Dikaryomycota originates; karyon is the Greek word for nut and in the lexicon of biology, it is the nucleus of a cell. The double nucleus dikaryon hypha can and does continue to grow by assimilation in a manner that is one of the most unusual aspects of fungal physiology. Starting with the first frame of the drawing, each of the two compatible nuclei divides by mitosis, retaining their haploid genetics so that two of each type is created – depicted in the second frame. One of the nuclei pairs separates so that one nucleus is at the tip of the hypha and the other, sister nucleus is at the back. The second nuclei pair also separates, but in this case one moves into a bulbous growth that protrudes from the side of the hypha while the other stays behind. As a new cell wall forms between the two sets of nuclei as shown in the third frame, the bulbous region clamps onto the hyphal wall on the other side of the septa, forming what is prosaically called a clamp connection. The clamp connection opens to allow the two separate nuclei to form an identical dikaryon and then closes so that at the end of the process in the fourth frame there is a dikaryon in each of the two separated cells. Laboratory observation has revealed that most basidiomycetes produce a new clamp connection about once every hour and that the mitotic division that creates the paired nuclei takes about 3 minutes. A sexually compatible dikaryon can continue to grow using clamp connections indefinitely as long as there is a nutritive source. Note that a 2n diploid has not yet been formed and meiosis, and therefore sex, has not yet occurred, just a lot of foreplay.

When environmental conditions are amenable for the successful dispersion of the sexually engendered spores, the reproductive cycle begins. It is the mechanics of the sexual process that differentiates the Subphylum Basidiomycotina from the Subphylum Ascomycotina; the former produce 4 spores on a structure called a basidium and the latter produce 8 spores on a structure called an ascus; the subphylum names are derived from the names of the two different spore containments. The conversion of the dikaryon into sexual spores by meiosis does not occur until the fungus is ready, a determination based on some means of determining temperature and moisture that registers the propensity for the spores to germinate if created. In the ascomycetes, the dikaryon undergoes a nuclear transition from n + n to the diploid 2n which rapidly undergoes meiosis to create four nuclei that divide by mitosis to create the 8 spores. The ascomycetes, which include the succulent edible morels and truffles, are mostly small, brightly colored cups like the Scarlet Cup (Sarcoscypha coccinea) or irregular stalks like the Orange Earth Tongue (Microglossum rufum); their taxonomy is established by the manner in which they release their spores from the ascus.

The basidiomyctes are the largest and most complex organisms of the Kingdom Eumycota; they are evident for their visible mushroom fruiting bodies. The reproductive machinations of basidiomycetes are essentially the same as those of the ascomycetes – a 2n diploid forms and divides by meiosis to create 4 haploid spores, stopping short of the final octal mitosis of the ascomycetes. The key difference is that in the basidiomycetes, a nascent, hypogeal reproductive fruiting body called the primordium is formed by the mycelium. It is only at the point of reproductive imminence that meiosis occurs; four club-shaped basidia are produced at the end of selected hyphae so that each of the four haploid spores will be separately housed and ready for dispersal. Fungi thus spend almost all of their lives as haploids contrasted to animals and plants that live almost wholly as diploids The spore producing surface is called the hymenium; for gilled mushrooms it is on both sides of the adjacent gills, for mushrooms with pores, it is the surface of the vertical tubes. The fully formed hypogeal mushroom primordium is now reproductively and physiologically ready to break the surface of the ground and open to expose its spore laden gills to the winds of chance dispersion. The impetus for this reproductive consummation is environmental. Mushrooms make their epigeal appearance soon after a rain as the conditions for spore germination are likely to be good. They also appear when their food source is threatened which may occur due to disease, death, fire, or, when that source is nutritionally depleted. If the spores germinate and find a mate, then fungal life goes on.
